2026年6月2日下午,首届霍夫曼讲坛(Hoffmann Lecture)在广州医科大学学术交流中心203报告厅隆重举行。广州霍夫曼免疫研究所荣誉所长兼首席科学家、2011年诺贝尔生理学或医学奖获得者Jules Hoffmann教授为首届霍夫曼讲坛致开幕辞。Hoffmann教授表示,他由衷欣喜地见证这项高水平学术活动启幕,这也标志着研究所的国际化发展翻开了崭新的重要篇章。过去十年来,广州霍夫曼免疫研究所始终保持蓬勃的学术活力,营造出稳定、开放、国际化的科研氛围。今天,我们创办霍夫曼讲坛,正是为了践行建所之初的使命。该讲坛将致力于打造长效、高水平的国际学术交流平台,汇聚全球顶尖科学家,分享前沿科研成果,碰撞创新学术思想,进一步提升研究所的国际影响力,推动全球免疫学与转化医学研究持续向前发展。

图1 霍夫曼教授致辞
霍夫曼讲坛的首场报告,我们荣幸地邀请到了赵金存教授。赵教授现任广州医科大学副校长、呼吸疾病全国重点实验室主任,是呼吸病毒学、新发传染病及免疫调控领域享誉业界的权威学者。
同呼吸,共命运。为何面对同一种呼吸道病毒感染,有人仅现轻微感冒症状,有人却发展为重症,甚至面临生命威胁?决定这场“免疫遭遇战”胜负的关键钥匙究竟藏在哪里?赵金存教授以题为“Interferon-stimulated gene GALNT2 restricts respiratory virus infections”的报告,为我们层层揭晓答案。

图2赵金存教授作报告与交流
冠状病毒(CoVs)与甲型流感病毒(IAVs)等呼吸道病毒,是引发全球大流行及人畜共患病传播的主要病原体,持续对人类公共卫生安全构成威胁。由SARS-CoV-2引发的新冠全球大流行,已造成超700万人死亡,对全球健康与社会经济发展带来深远影响;2009年甲型流感疫情作为21世纪首次全球性流感大流行,也导致了至少20万人死亡。
在轻症感染情况下,机体能够快速启动正常且高效的免疫应答反应:干扰素(IFNs)及时激活干扰素刺激基因(ISGs),快速建立抗病毒防御屏障,有效抑制病毒早期复制并限制其扩散。相比之下,重症感染常伴随免疫应答延迟、功能缺陷或被抑制等特征。部分患者因干扰素系统存在有害的等位基因变异,或体内产生干扰素中和抗体,直接削弱先天抗病毒能力;机体无法及时清除病毒,导致病毒持续增殖与扩散,最终彻底打破免疫平衡。
已有研究证实,由IFNs诱导表达的单个ISG能够有效增强机体的抗病毒防御能力,显著降低COVID-19重症发生风险,这表明ISG介导的抗病毒应答是机体抵御呼吸道病毒侵袭、缓解重症感染的核心保护性机制。然而,相较于对I型干扰素(IFN-I)整体通路的宏观研究,单个ISG的特异性抗病毒功能及分子调控机制仍未被完全阐明。因此,系统挖掘特定ISG在抑制冠状病毒、甲型流感病毒感染过程中的关键作用与调控机理,对于解析先天免疫防御机制、开发广谱抗病毒靶点,具有重要的科学意义与应用价值。
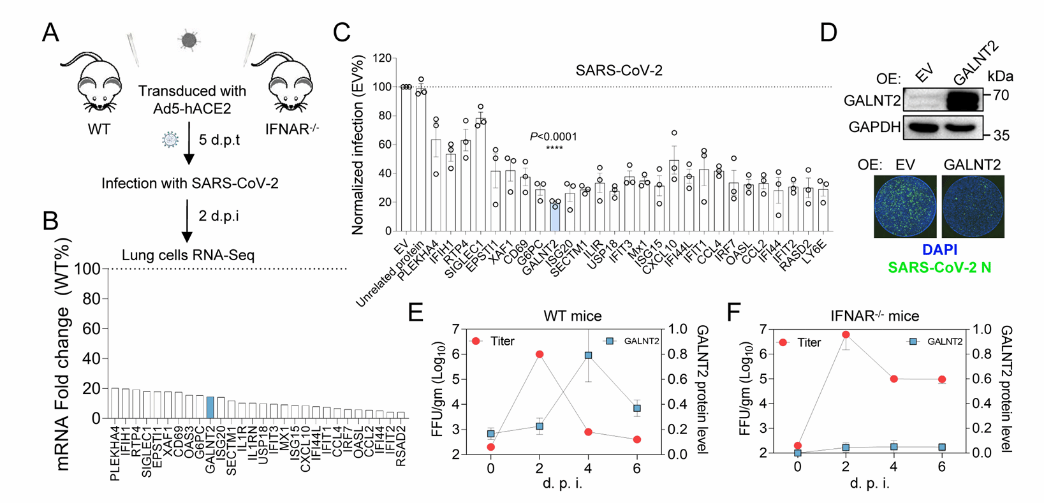
图3 筛选策略
筛选并鉴定与呼吸道病毒感染严重程度相关的保护性ISG:GALNT2
传统筛选策略通常通过感染病毒后检测基因转录水平的变化来识别关键基因。赵老师团队则另辟蹊径,对免疫缺陷小鼠,即干扰素α/β受体敲除(IFNAR−/−)小鼠,进行SARS-CoV-2感染,寻找感染后表达下调的基因,从而更精准地锚定干扰素诱导信号通路。通过这一策略,他们发现了28个明显下调的ISG(图3-A, B)。随后,团队在Huh7细胞中过表达这些候选基因,并开展SARS-CoV-2感染实验,发现一种编码糖基转移酶的基因(GALNT2)能够有效降低病毒的增殖水平(图3-C, D)。在野生型小鼠感染SARS-CoV-2后,肺部GALNT2蛋白表达量自感染后第2天起迅速上升,于第4天达到峰值。而在免疫缺陷小鼠中,GALNT2蛋白水平降低与病毒载量升高之间存在相关性,提示GALNT2表达受抑可能导致新冠病毒持续复制(图3-E, F)。
为进一步探究GALNT2在急性重症新冠患者中的表达情况,赵老师团队对患者的支气管肺泡灌洗液及外周血单核细胞样本进行了RNA测序。结果显示,新冠患者体内GALNT2表达水平高于健康对照,且轻症患者高于重症患者。通过对不同感染阶段(T1:初始感染期;T2:转化期;T3:恢复期)免疫细胞的单细胞转录组测序追踪,发现轻症患者的GALNT2表达水平在T2阶段显著升高(图4-A-D)。这些发现表明,GALNT2在促进病毒清除、减轻疾病严重程度及加速康复过程中发挥重要作用。全基因组关联分析(GWAS)进一步揭示,GALNT2基因存在单核苷酸多态性(SNPs),与COVID-19及IAV(包括H1N1和H7N9)感染的易感性、疾病严重程度及住院风险升高显著相关(图4-E, F)。
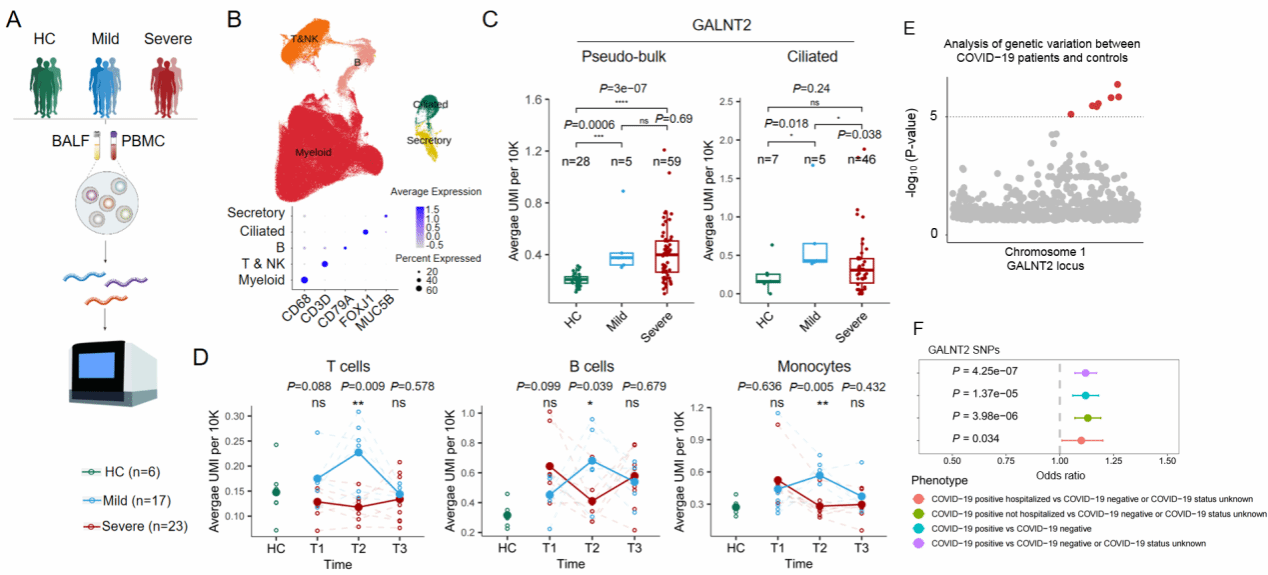
图4 GALNT2 在新冠疾病发展中的变化情况
GALNT2 是一种具有广谱抗病毒活性的 ISG
为深入探究GALNT2的抗病毒功能,赵老师团队首先在Huh7细胞中过表达GALNT2,发现相较于对照组,GALNT2在不同时间点均显著抑制了SARS-CoV-2(野生型及Omicron变异株)的复制(图5-A)。为进一步明确GALNT2是否具备广谱抗病毒活性,团队在Huh7细胞和A549细胞中分别感染了多种冠状病毒(CoVs)。结果显示,GALNT2的过表达显著抑制了冠状病毒及甲型流感病毒的复制(图5-A, B)。相反,利用CRISPR-Cas9技术构建的Huh7(GALNT2−/−)细胞中,内源性GALNT2的敲除显著促进了SARS-CoV-2的复制(图5-C)。值得注意的是,GALNT2−/−细胞对病毒复制的敏感性与STAT1−/−细胞相似(图5-C),提示GALNT2是一个关键的抗病毒效应因子。此外,在敲除细胞中重新回补GALNT2表达后,其抗病毒活性得以恢复(图5-C),表明内源性与外源性GALNT2均能广谱抑制新冠病毒和甲型流感病毒的复制。
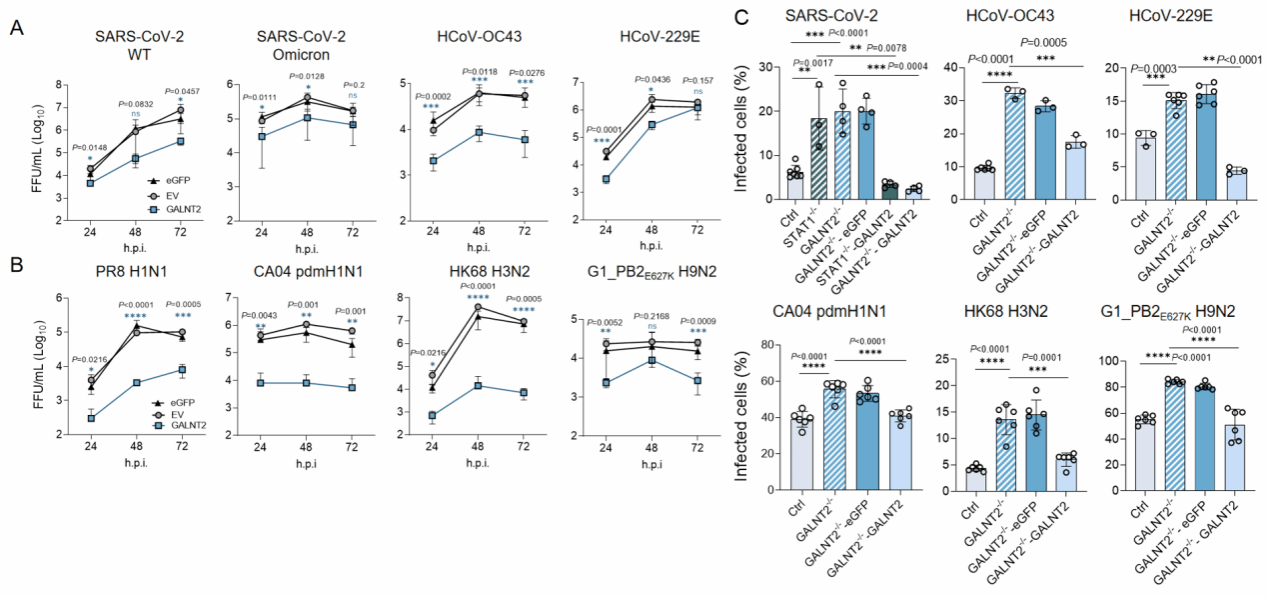
图5 GALNT2的广泛抗病毒效果
GALNT2 体内保护小鼠抵御 SARS-CoV-2 和 IAV 感染
接下来,赵老师团队进一步探究了GALNT2在体内的抗病毒效果。他们利用CRISPR-Cas9基因编辑技术构建了内源性GALNT2基因敲除小鼠(GALNT2−/−)。用亚致死剂量的PR8毒株感染后,GALNT2−/−小鼠相较于野生型(WT)小鼠,体重减轻更显著、存活率更低、肺部病毒载量更高,且肺组织病理损伤更严重(图6-A),表明GALNT2对甲型流感病毒(IAV)感染具有保护作用。此外,在SARS-CoV-2感染实验中,GALNT2−/−小鼠肺部的病毒载量也高于WT小鼠,尽管两者体重变化无显著差异(图6-B)。这些结果共同表明,GALNT2在体内具有强大的抗病毒活性。
进一步,团队采用血清9型腺相关病毒(AAV9)作为基因转移载体,通过鼻腔途径向WT小鼠递送AAV9-GALNT2。结果发现,在PR8或SARS-CoV-2致死剂量攻击下,与空载体(EV)对照组相比,50%的GALNT2转导小鼠存活,且仅表现出轻微的体重下降和较低的肺部病毒载量(图6-C, D)。此外,团队还探究了GALNT2能否保护免疫缺陷小鼠免受SARS-CoV-2感染。为此,他们用SARS-CoV-2感染了经GALNT2转导的IFNAR−/−和STAT1−/−小鼠,结果发现肺部病毒滴度降低了约10倍(图6-E),进一步支持了GALNT2在免疫缺陷个体中的抗病毒潜力。
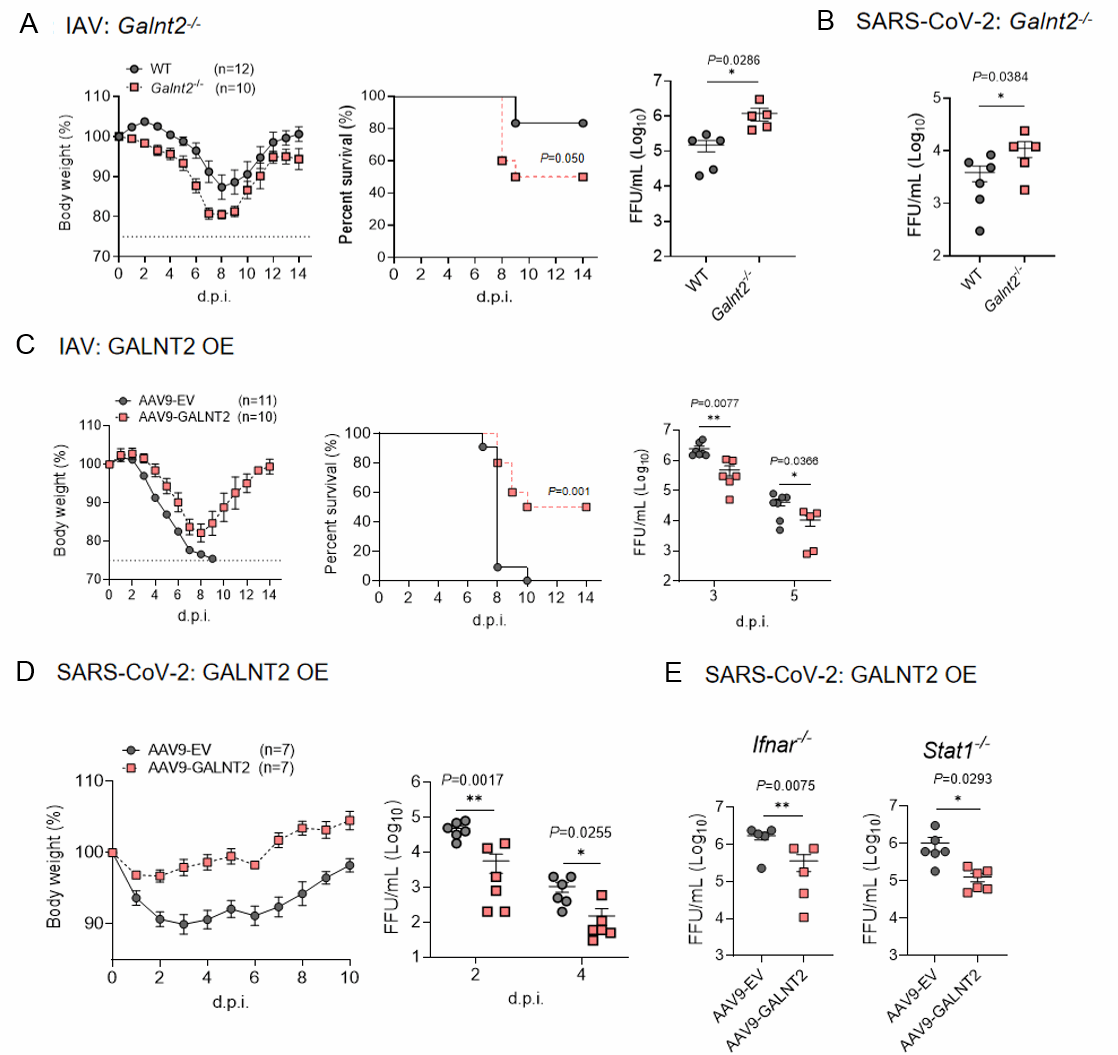
图6 GALNT2 在小鼠模型中抑制 SARS-CoV-2 和 IAV 的体内复制
GALNT2 特异性 O-糖基化修饰刺突蛋白 S2′切割位点上游残基
GALNT2 是 20 种 N-乙酰半乳糖胺基转移酶(GALNTs)之一,负责启动特定蛋白底物的 O-糖基化修饰。赵老师团队通过组学数据发现,GALNT2 过表达在 RNA 和蛋白质水平上对多数宿主基因的表达水平和宿主蛋白的O-糖基化水平影响甚微,提示GALNT2 依赖性的 O-糖基化可能直接修饰病毒蛋白。SARS-CoV-2 刺突蛋白在病毒组装过程中首先被前蛋白转化酶Furin蛋白酶在 S1/S2 切割;随后,TMPRSS2在病毒进入及细胞-细胞融合(合胞体形成)过程中对刺突蛋白的 S2′位点进行二次切割。团队观察到,在GALNT2过表达的细胞中产生的子代病毒,其形成的空斑体积显著减小,据此推测GALNT2可能通过阻断刺突蛋白介导的病毒-细胞融合而发挥功能。
为验证该假说,研究者利用体外糖基化实验检测GALNT2对包含蛋白酶切割位点的合成肽段的酶活性,结果显示GALNT2可特异性糖基化TMPRSS2(S2′)切割位点附近的肽段序列,而对Furin(S1/S2)位点无作用。液相色谱-串联质谱(LC–MS/MS)进一步鉴定出,GALNT2特异性修饰野生型SARS-CoV-2刺突蛋白中对应于丝氨酸810和813(S810/813)的残基(图7-A-C)。随后,他们对过表达的刺突蛋白胞外域(ECD)进行了基于LC–MS的糖谱分析,并证实体外糖基化残基中至少有一个(S813)存在于重组刺突蛋白上(图7-D)。这些结果明确了GALNT2对刺突蛋白S2′切割位点邻近残基的直接修饰作用,为理解其抗病毒机制提供了分子层面的证据。
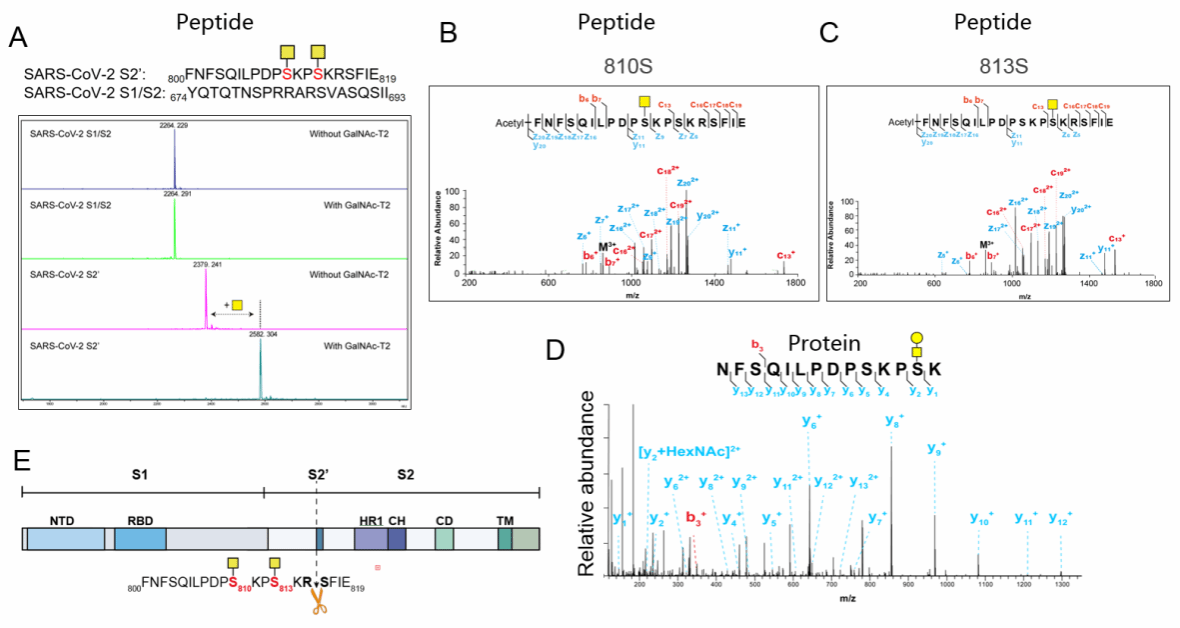
图7 GALNT2 特异性 O-糖基化修饰刺突蛋白 S2′切割位点上游的残基
GALNT2通过阻断病毒与细胞融合来抑制病毒生长
随后,赵老师团队进一步探究了GALNT2介导的修饰是否影响S2′位点的切割。在共表达刺突蛋白与ACE2的细胞中,可清晰检测到S2′切割产物;然而,与对照组相比,在HEK293T细胞(图8-A)及HEK293T-TMPRSS2细胞(图8-B)中,共表达GALNT2均显著减少了S2′切割。为排除GALNT2对上游S1/S2 Furin切割位点的潜在间接影响,他们检测了S1/S2连接处缺失ΔRRAR的刺突突变体。结果显示,GALNT2仍能有效减少该突变体的S2′切割(图8-C),证实GALNT2是通过对刺突蛋白S810/S813位点的O-糖基化修饰,直接抑制其蛋白水解活化。鉴于GALNT2可抑制病毒糖蛋白的蛋白水解切割,研究者推测其可能削弱切割依赖性的膜融合能力。为评估GALNT2存在时刺突蛋白的促融合活性,他们利用Cre-loxP报告系统进行了定性与定量的细胞-细胞融合实验。共表达刺突蛋白与GALNT2时,两种信号均降低约50%(图8-D),表明刺突蛋白介导的膜融合受到显著抑制。进一步的功能验证显示,GALNT2过表达不仅降低了SARS-CoV-2假病毒及病毒样颗粒的进入效率,还显著削弱了多种高致病性流感病毒HA蛋白介导的膜融合活性。这些发现表明,GALNT2 介导的病毒表面糖蛋白的 O-糖基化限制了膜融合并抑制了病毒进入。
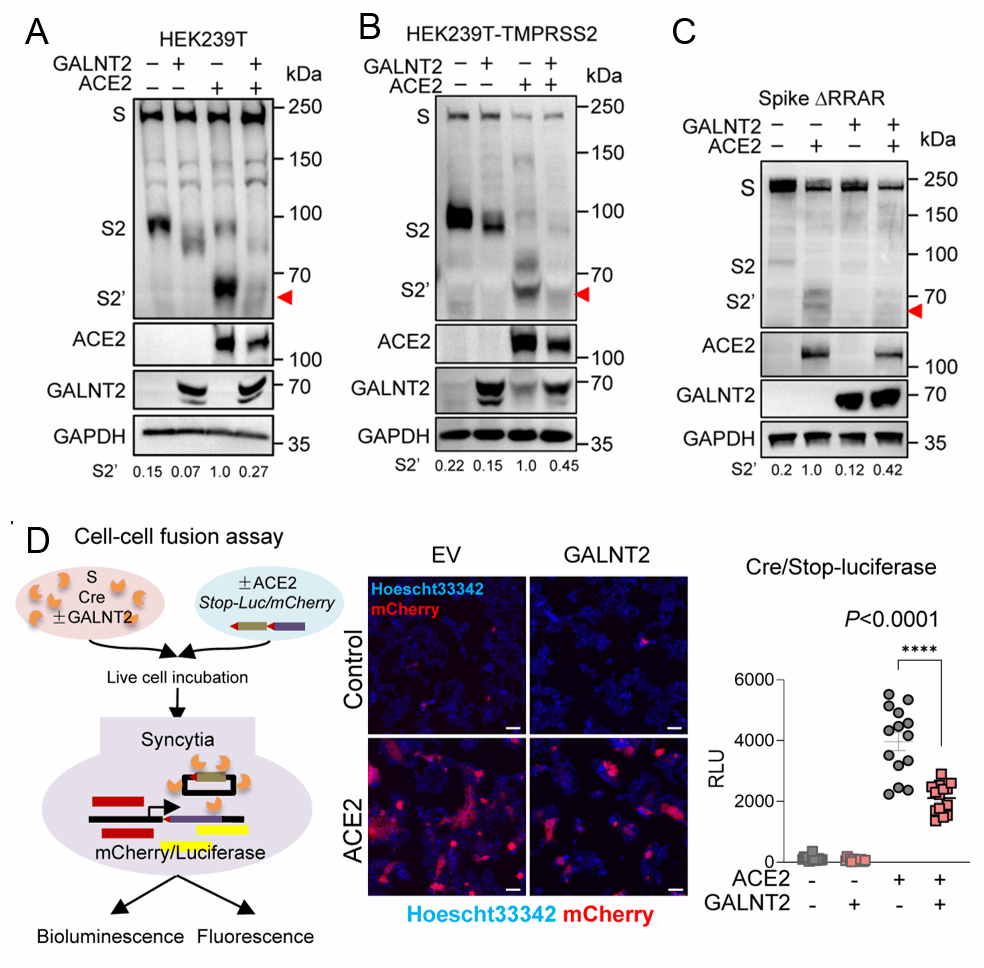
图8 GALNT2通过阻断病毒与细胞融合来抑制病毒生长
GALNT2抗病毒关键靶点的验证
接下来,赵老师团队进一步明确SARS-CoV-2刺突蛋白S810/S813位点是否为GALNT2发挥抗病毒功能的主要靶标。通过序列追踪,他们发现SARS-CoV-2 B.1.36.25分支在2020年底前后曾短暂出现过I813天然替代突变,但该变异株很快消失(图9-A),提示该突变可能伴随适应性代价,使得病毒虽能抵抗GALNT2修饰,却难以在人群中长期维持。为排除S810位糖基化的潜在影响,研究者分别在刺突蛋白中引入S810/813I和S810/813A双点突变,并制备了相应突变体的重组病毒(图9-B)。结果显示,与野生型病毒相比,这两种重组病毒在Vero E6及Huh7细胞中的病毒滴度及空斑直径均显著降低(图9-C-E)。同时,二者在体外和体内均表现出对GALNT2限制效应的显著抵抗(图9-F,G)。这些结果表明,刺突蛋白S810/S813位点在决定病毒对GALNT2的敏感性或抵抗性中起着关键作用。
随后,研究者将S810/813I突变引入可视化病毒复制子,结果发现该突变复制子对GALNT2的抗病毒活性表现出严格抵抗。此外,GALNT2过表达能明显抑制野生型SARS-CoV-2诱导的合胞体形成,但对S810/813I突变复制子则无显著影响。结果,赵老师团队认为,GALNT2介导的O-糖基化修饰构成了一种高度保守的天然免疫策略,病毒若试图通过点突变逃避该修饰,则需付出复制适应性和传播能力的显著代价,这使得GALNT2介导的抗病毒通路难以被病毒轻易突破,为抗病毒干预提供了潜在且稳健的靶点。
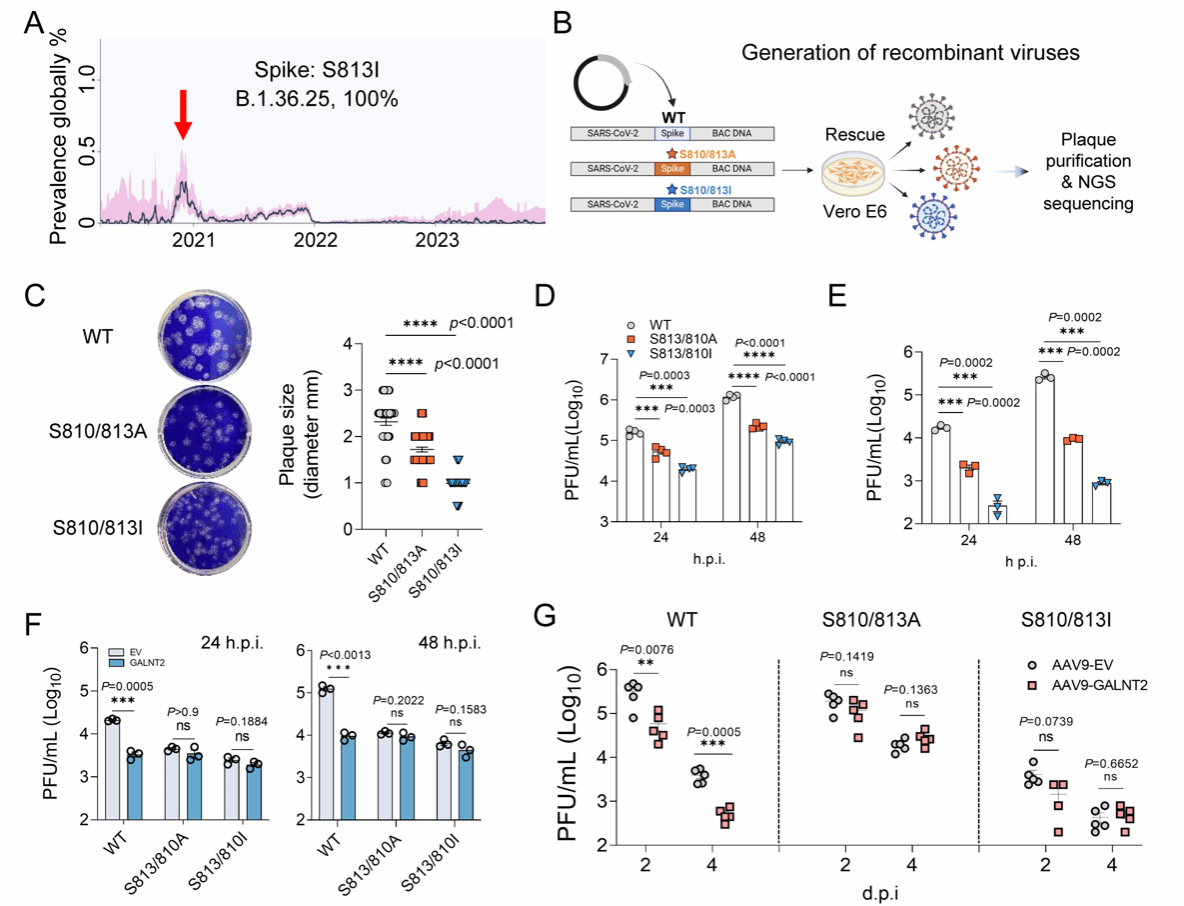
图9 GALNT2靶向刺突蛋白S810/S813:病毒逃逸受限的天然免疫机制
从逆向思维到科研之道
在交流互动环节,与会师生对赵老师团队独特的“逆向思维”研究路径表现出浓厚兴趣。他们从敲除小鼠出发,进行转录组测序,重点关注表达下调的基因,并从28个候选者中筛选出尚未被报道的分子展开深入研究,最终取得突破性成果。
针对部分研究生在课题方向上的困惑,赵老师表示,科研能力和兴趣都是可以后天培养的。他介绍道,实验室的学生通常同时承担两个课题,在相互协作中共同成长。关于“什么样的人适合做科研”,赵老师认为,潜力是最重要的素质,一个人唯有具备可塑性,才能在科研道路上走得长远。
报告结束后,Jean-Luc Imler教授代表研究所向赵金存教授赠送了纪念品(图10)。我们再次对赵教授带来的精彩报告致以诚挚感谢!

图10 为赵金存教授颁发霍夫曼讲坛纪念奖章
(左起)Jules Hoffmann教授、赵金存教授、Jean-Luc Imler教授、焦仁杰教授
参考文献
[1] Ran, W., Yang, J., Yu, S. et al. Interferon-stimulated gene GALNT2 restricts respiratory virus infections. Nat Microbiol., 2026, 11, 256–270.
